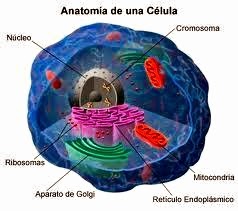
El citoplasma: funciones y orgánulos
El objetivo de este artículo es describir la estructura y las funciones del citoplasma y los orgánulos. El citoplasma está constituido por todos los elementos que se hallan por dentro de la membrana plasmática, excepto el núcleo, y tiene dos componentes que son el citosol y los orgánulos, pequeñas estructuras que realizan distintas funciones en la célula. Esta parte de las células cumple importantes funciones como por ejemplo producir muchas enzimas que ayudan a catalizar la glucólisis (degradación de glucógeno), lo cual se lleva a cabo en el citosol.
El citosol (líquido intracelular) es la porción líquida del citoplasma que rodea a los orgánulos y constituye alrededor del 55% del volumen total. A pesar que varía en su composición y consistencia en los distintos sectores de la célula, el citosol está formado en un 70 a 95% por agua, a la que se suman diferentes compuestos que están disueltos o en suspensión.
Entre éstos hay diferentes tipos de iones, glucosa, aminoácidos, ácidos grasos, proteínas, lípidos, ATP y productos de desecho. Ciertas células también presentan diversas moléculas orgánicas que se almacenan en cúmulos. Estos agregados pueden aparecer y desaparecer en diferentes momentos de la vida de una célula. Algunos ejemplos son las "gotas de lípido" que contienen trglicéridos y las partículas de moléculas de glucógeno denominadas gránulos de glucógeno.
El citosol es el lugar donde se producen muchas de las reacciones químicas necesarias para mantener viva a la célula. Por ejemplo, las enzimas del citosol catalizan la glucólisis, una serie de diez reacciones químicas que dan lugar a dos moléculas de ATP a partir de una molécula de glucosa. Otros tipos de reacciones citosólicas aportan los materiales de construcción fundamentales para el mantenimiento y crecimiento de las estructuras celulares.
Los orgánulos son estructuras especializadas dentro de la célula que tienen unas formas características y que llevan a cabo funciones específicas en el crecimiento, mantenimiento y reproducción celulares. A pesar de la diversidad de reacciones químicas que tienen lugar en una célula en un momento dado, existe muy poca interferencia entre las reacciones ya que están confinadas en diferentes orgánulos.
Cada uno de ellos tiene su propio grupo de enzimas que llevan a cabo reacciones específicas y funcionan como unidades compartimentales para estos procesos bioquímicos determinados. El número y el tipo de orgánulos varían en las diferentes células, de acuerdo con la función que cumplen. A pesar de tener diferentes funciones, los orgánulos cooperan unos con otros para mantener la homeostasis. Si bien el núcleo es un orgánulo grande, será descrito en este artículo debido a su especial importancia en e control del ciclo vital de las células.

El citoesqueleto es una red de filamentos proteicos que se extienden a través del citosol. Tres tipos de filamentos proteicos contribuyen a la estructura del citoesqueleto, así como la de otros orgánulos. En orden creciente de diámetro, estas estructuras son los microfilamentos, los filamentos intermedios y los microtúbulos.
Los microfilamentos, los elementos más delicados del citoesqueleto, están compuestos por la proteína actina y son más abundantes en la periferia de la célula. Tienen dos funciones generales: ayudan a generar el movimiento y proveen soporte mecánico. Con respecto al movimiento, los microfilamentos intervienen en la contracción muscular, la división y la locomoción celulares, como la que se produce en la migración de las células embrionarias durante el desarrollo, la invasión de diferentes tejidos por los glóbulos blancos (leucocitos) para combatir la infección o la migración de células cutáneas en el proceso de curación de las heridas.
Los microfilamentos proporcionan la mayor parte del soporte mecánico que es responsable de la fuerza y la forma básicas de la célula. Estos microfilamentos unen el citoesqueleto a las proteínas integrales de la membrana plasmática, también dan soporte mecánico a las extensiones celulares denominadas microvellosidades, proyecciones en forma de dedo, inmóviles, de la membrana plasmática. Dentro de cada microvellosidad hay un núcleo de microfilamentos paralelos. Puesto que aumentan en gran medida la superficie celular, las microvellosidades son abundantes en las células que están relacionadas con la absorción, como las células epiteliales del intestino delgado.
Como su nombre lo sugiere, los filamentos intermedios son más gruesos que los microfilamentos pero más delgados que los microtúbulos. Varios tipos de proteínas pueden formar parte de los filamentos intermedios, que son excepcionalmente fuertes. Se encuentran en las regiones de la célula sujetas a estrés mecánico, ayudan a estabilizar la ubicación de orgánulos como el núcleo, y contribuyen a que las células se adhieran entre sí.
El componente más grande del citoesqueleto, los microtúbulos, son tubos largos, huecos y no ramificados, compuestos principalmente por la proteína tubulina. El ensamblado de los microtúbulos comienza en un orgánulo llamado centrosoma. Los microtúbulos se desarrollan desde los centrosomas en la periferia de la célula, y contribuyen a dar forma a la célula. También cumplen funciones en el movimiento de ciertos orgánulos como las vesículas secretoras, de los cromosomas durante la división celular y de las proyecciones celulares especializadas, como los cilios y los flagelos.

El centrosoma, localizado cerca del núcleo, tiene dos componentes: un par de centríolos y material pericentriolar. El centríolo es una estructura cilíndrica compuesta por nueve complejos de tres microtúbulos (tripletes) dispuestos en forma circular. El eje longitudinal de uno de los centríolos está en ángulo recto con el eje longitudinal del otro. Alrededor de los centríolos se encuentra el material centriolar, que contiene cientos de complejos anulares formados por la proteína tubulina. Estos complejos de tubulina son los centros de organización del crecimiento del huso mitótico, que desempeña un papel crítico en la división celular y también en la formación de microtúbulos en las células que no están en división activa. Durante la división celular, los centrosomas se replican de manera que las generaciones sucesivas de células tengan la capacidad de realizar la división celular

Los microtúbulos con los componentes predominantes de los cilios y flagelos, protecciones móviles de la superficie celular. Los cilios son apéndices numerosos, cortos, similares a pelos, que se extienden desde la superficie de la célula. Cada cilio contiene un núcleo de 20 microtúbulos rodeado por la membrana plasmática. Los microtúbulos están dispuestos de tal manera que un par central es rodeado por nueve complejos de dos microtúbulos fusionados (dobletes). Cada cilio permanece unido a un cuerpo basal que se halla por debajo de la superficie de la membrana plasmática. Un cuerpo basal es similar en su estructura a un centríolo y cumple funciones en el ensamblado de los cilios y flagelos.
Los cilios realizan un movimiento similar al de un remo; es relativamente rígido durante el movimiento de fuerza (el remo introduciéndose en el agua), pero es más flexible durante el movimiento de recuperación (el remo moviéndose por arriba del agua y preparándose para una nueva remada). Los movimientos coordinados de muchos cilios en la superficie de una célula dan lugar a un movimiento sostenido del líquido a lo largo de la superficie celular.

Muchas células del tracto respiratorio, por ejemplo, tiene cientos de cilios que ayudan a barrer fuera de los pulmones las partículas extrañas atrapadas en el moco. En la fibrosis quística, la densidad anormal de las secreciones mucosas interfiere con la acción de los cilios y, por ende, con las funciones normales del tracto respiratorio. El movimiento de los cilios también es paralizado por la nicotina del humo del cigarrillo. Por esta razón, los fumadores tosen con frecuencia para eliminar las partículas extrañas de sus vías respiratorias. Las células que revisten las trompas uterinas (de Falopio) también tienen cilios que desplazan al óvulo hacia el útero, y las mujeres que fuman tienen un riesgo mayor de embarazo ectópico (fuera del útero).
Los flagelos son similares en su estructura a los cilios, pero suelen ser mucho más largos. Los flagelos generalmente mueven una célula entera. In flagelo genera movimiento hacia adelante a lo largo de su eje por el movimiento rápido que sigue un patrón ondulante. El único ejemplo de flagelos en el cuerpo humano e la cola de los espermatozoides, que propulsa a estas células hacia su encuentro con el óvulo.

Los ribosomas son los sitios de síntesis proteica. El nombre de estos pequeños orgánulos refleja su alto contenido de un tipo especial de ácido ribonucleico, el ARN ribosómico (ARNr), aunque también puede incluir a más de 50 proteínas. Estructuralmente, un ribosoma está constituido por subunidades, una de las cuales tiene la mitad del tamaño de la otra. Las subunidades mayor y menor se forman por separado en el nucléolo, un cuerpo esférico que está dentro del núcleo. Una vez sintetizadas, las subunidades mayor y menor abandonan el núcleo y se unen en el citoplasma.
Algunos ribosomas están adheridos a la superficie externa de la membrana nuclear y a una membrana extensamente plegada denominada retículo endoplasmático. Estos ribosomas sintetizan las proteínas que tienen como destino orgánulos específicos, su inserción en la membrana plasmática o su salida de la célula. Otros ribosomas son "libres" o no adheridos a ninguna otra estructura citoplasmática. Los ribosomas libres sintetizan proteínas que serán utilizadas en el citosol. Los ribosomas también están localizados dentro de las mitocondrias, donde sintetizan proteínas mitocondriales.

El retículo endoplsamático es una red de membranas en forma de sacos aplanados o túbulos. El retículo endoplasmático se extiende desde la membrana o envoltura nuclear (membrana que rodea al núcleo), con la cual se conecta hacia todo el citoplasma. El retículo endoplasmático es tan amplio que constituye casi la mitad de las superficies membranosas dentro del citoplasma de la mayoría de las células.
Las células contienen dos tipos distintos de retículo endoplasmático, que difieren tanto en su estructura como en su función. El retículo endoplasmático rugoso es una estructura que continúa con la membrana nuclear y generalmente se presenta plegado a una serie de sacos aplanados. La superficie externa del retículo endoplasmático rugoso está revestida de ribosomas, los sitios de síntesis proteica. Las proteínas sintetizadas por los ribosomas penetran en los espacios que hay en el interior del retículo endoplasmático rugoso para su procesamiento y distribución. En algunos casos, ciertas enzimas unen proteínas con hidratos de carbono para formar glucoproteínas. En otros casos, las enzimas unen proteínas con fosfolípidos, también sintetizados en el retículo endoplasmático rugoso. Estás moléculas pueden incorporarse a las membranas de los orgánulos, insertarse en la membrana plasmática o secretarse por exocitosis. De tal modo que el retículo endoplasmático rugoso produce proteínas secretoras, proteínas de membrana y muchas proteínas de los orgánulos.
El retículo endoplasmático liso se extiende desde el retículo endoplasmático rugoso para formar una red de túbulos membranosos. A diferencia del R.E. rugoso, el retículo endoplasmático liso carece de ribosomas en la superficie externa de sus membranas. Sin embargo, contiene enzimas especiales que pueden hacerlo funcionalmente más diverso que el retículo endoplasmático rugoso. La ausencia de ribosomas impide la síntesis de proteínas, pero sí puede sintetizar ácidos grasos y esteroides, como los estrógenos y la testosterona. En las células hepáticas, las enzimas del retículo endoplasmático liso facilitan la liberación de la glucosa hacia el torrente sanguíneo y contribuyen a inactivar o detoxificar los fármacos liposolubles o las sustancias potencialmente nocivas, como el alcohol, los pesticidas y los carcinógenos (agentes causantes de cáncer). En las células del hígado, del riñón y del intestino, una enzima del retículo endoplasmático liso separa el grupo fosfato de la glucosa-6-fosfato, lo cual permite que se libere la glucosa y pueda ingresar en la sangre. En las células musculares, los iones calcio que estimulan la contracción son liberados desde el retículo sarcoplasmático, una forma del retículo endoplasmático liso.
Una de las funciones del retículo endoplasmático liso es detoxificar ciertos fármacos. Quienes consumen de forma repetido algunos fármacos, como el sedante fenobarbital, desarrollan cambios en el retículo endoplasmático listo de las células hepáticas. La administración prolongada de fenobarbital trae como resultado la tolerancia al fármaco; la misma dosis deja de producir el mismo grado de sedación. Con la exposición reiterada al fármaco, la cantidad del retículo endoplasmático liso y sus enzimas aumentan para proteger a la célula de sus efectos tóxicos. Como la cantidad de retículo endoplasmático liso aumenta, son necesarias dosis cada vez más altas del fármaco para alcanzar el grado original del efecto.

La mayor parte de las proteínas sintetizadas en los ribosomas adheridos al retículo endoplasmático rugoso son por último transportadas a otras regiones de la célula. El primer paso en la vía de transporte es un orgánulo denominado complejo de Golgi. Consta de 2 a 20 cisternas, pequeños sacos aplanados membranosos, de bordes salientes, que parecen una pila de pitas (pan árabe). Las cisternas suelen ser curvadas y le dan forma al complejo de Golgi un aspecto de copa. Casi todas las células tienen varios complejos de Golgi, y éstos son más numerosos en las células que secretan proteínas, una clave para entender el papel de este orgánulo en la célula.
Las cisternas en los extremos opuestos de un complejo de Golgi difieren entre sí en su forma, su tamaño y su actividad enzimática. La entrada convexa o cara cis, mira hacia el retículo endoplasmático rugoso. La salida cóncava o cara trans, mira hacia la membrana plasmática. Los sacos que se hallan entre las caras de entrada y de salida se denominan cisternas mediales. Las vesículas de transporte provenientes del retículo endoplasmático se fusionan para formar la cara de entrada. Se piensa que las cisternas mediales y luego en cisternas de salida.
Las diferentes enzimas presentes en las regiones de entrada, medial y de salida del complejo de Golgi permiten que cada una de estas áreas pueda modificar, ordenar y envolver a las proteínas para su transporte a diferentes destinos. La cara de entrada recibe y modifica las proteínas producidas por el retículo endoplasmático rugoso. Las cisternas mediales agregan hidratos de carbono a las proteínas para formar glucoproteínas y lípidos para formar lipoproteínas. La cara de salida modifica aún más las moléculas y luego las selecciona y reúne para el transporte hacia su destino final.
Las proteínas que llegan al complejo de Golgi, lo atraviesan y salen de éste mediante la maduración de las cisternas y los intercambios que ocurren por las vesículas de transferencia.

1. Las proteínas sintetizadas por los ribosomas del retículo endoplasmático rugoso son rodeadas por una porción de la membrana del retículo endoplasmático, que posteriormente brota de la superficie de la membrana para formar las vesículas de transporte.
2. Las vesículas de transporte se dirigen hacia la cara de entrada del complejo de Golgi.
3. La fusión de varias vesículas de transporte crea una cara de entrada en el complejo de Golgi y libera proteínas dentro de su luz (espacio).
4. Las proteínas se mueven desde la cara de entrada hacia una o más cisternas mediales. Las enzimas de estas cisternas modifican las proteínas para formar glucoproteínas, glucolípidos, y lipoproteínas. Las vesículas de transferencia que surgen de los bordes de las cisternas llevan enzimas específicas de regreso a la cara de entrada y transportan algunas proteínas parcialmente modificadas hacia la cara de salida.
5. Los productos de las cisternas mediales se mueven hacia la luz de la cara de salida.
6. Dentro de la cara de salida, los productos sufren más modificaciones, se clasifican y rodean de membrana y son empaquetados.
7. Algunas de las proteínas procesadas abandonan la cara de saluda y quedan almacenadas en vesículas secretoras. Estas vesículas llevan las proteínas hacia la membrana plasmática, donde se liberan por exocitosis hacia el líquido extracelular. Ciertas células pancreáticas, por ejemplo, liberan la hormona insulina de esta forma.
8. Otras proteínas procesadas abandonan la cara de salida en vesículas de membrana que entregan su contenido a la membrana plasmática para su incorporación dentro de la propia membrana. De esta manera, el complejo de Golgi agrega unos nuevos segmentos a la membrana plasmática a medida que los preexistentes se pierden y modifica el número y la distribución de las moléculas de la membrana.
9. Finalmente, algunas proteínas procesadas abandonan la cara de salida de las vesículas de transporte que las llevan hacia otro destino en la célula. Por ejemplo, las vesículas de transporte llevan las enzimas digestivas hacia los lisosomas.

Los lisosomas son vesículas rodeadas de membrana que se forman en el complejo de Golgi. En su interior hay más de 60 tipos de poderosas enzimas digestivas e hidrolíticas que pueden digerir una gran variedad de moléculas una vez que los lisosomas se fusionaron con las vesículas formadas durante la endocitosis. Como las enzimas lisosómicas funcionan mejor en un pH ácido, la membrana lisosómica contiene bombas de transporte activas que importan iones hidrógeno (H+). De esta manera, el interior de los lisosomas tiene un pH de 5, que es 100 veces más ácido que el pH del citosol (pH 7). La membrana lisosómica también presenta transportadores que acarrean los productos finales de la digestión, como la glucosa, los ácidos grasos y los aminoácidos, hacia el citosol.
Las enzimas lisosómicas intervienen, así mismo, en el reciclado de las estructuras celulares deterioradas. Un lisosoma puede ingerir otro orgánulo, digerirlo y luego devolver los componentes digeridos al citosol para su nuevo uso, proceso que se denomina autofagia. Durante la autofagia, el orgánulo que ha de ser digerido es rodeado por una membrana que deriva del retículo endoplasmático para crear la vesícula denominada autofagosoma, que luego se fusiona con un lisosoma. De tal modo, una célula hepática humana recicla alrededor de la mitad de su contenido citoplasmático cada semana. Las enzimas lisosómicas también pueden destruir la célula entera mediante el proceso de autólisis. La autólisis tiene lugar en algunas situaciones patológicas y es también responsable del deterioro de los tejidos después de la muerte.
La mayor parte de las enzimas lisosómicas actúan dentro de la célula. Sin embargo, algunas participan en la digestión extracelular. Un ejemplo se da durante la fecundación. La cabeza del espermatozoide libera enzimas lisosómicas que lo ayudan a introducirse en el óvulo, por la disolución de su cubierta protectora en el proceso llamado reacción acrosómica.
Otro grupo de orgánulos similares en su estructura a los lisosomas, pero más pequeños, son los peroxisomas. Contienen varias oxidasas, enzimas que pueden oxidar (eliminar átomos de hidrógeno) de diversas sustancias orgánicas. Por ejemplo, los aminoácidos y los ácidos grasos pueden ser oxidados en los peroxisomas como parte del metabolismo normal. Además, las enzimas de los peroxisomas oxidan sustancias tóxicas, como el alcohol. Por ello son muy abundantes en el hígado, donde tiene lugar la detoxificación del alcohol y otras sustancias nocivas. Un producto intermedio de las reacciones de oxidación es el peróxido de hidrógeno (H2O2), un compuesto potencialmente tóxico. Sin embargo, los peroxisomas también contienen la enzima catalasa, que descompone al H2O2. Como la producción y la degradación del H2O2 tiene lugar dentro de la misma organela, los peroxisomas protegen a otras partes de la célula de los efectos tóxicos del H2O2. Los peroxisomas nuevos se forman a partir de los ya existentes.
Como ya mencionó, los lisosomas degradan las proteínas que reciben por medio de las vesículas. Las proteínas citosólicas también requieren su eliminación en algún momento del ciclo vital de una célula. La destrucción permanente de las proteínas no necesarias, dañadas o defectuosas es una función que está a cargo de pequeñas estructuras con forma de tonel denominadas proteasomas. Las proteínas que forman parte de las vías metabólicas, por ejemplo, son degradas después de haber cumplido su función. Esta destrucción proteica participa en la retroalimentación negativa ya que establece un alto en la vía una vez que se obtuvo la respuesta adecuada. Una célula típica del organismo contiene muchos cientos de proteasomas, tanto en el citosol como en el núcleo. Los proteasomas reciben su nombre debido a su alto contenido de proteasas, enzimas que pueden cortar a las proteínas en péptidos pequeños. Una vez que las enzimas de un proteasoma hidrolizaron a una proteína en segmentos más pequeños, otras enzimas pueden luego descomponer a esos péptidos en aminoácidos, los cuales se reciclan para formar nuevas proteínas.
Algunas enfermedades son el resultado de la incapacidad de los proteasomas de degradar proteínas anormales. Por ejemplo, se acumulan grupos de proteínas con defectos de plegamiento en las células cerebrales en los pacientes con enfermedad de Parkinson y enfermedad de Alzheimer. El descubrimiento de por qué los proteasomas no pueden degradar a estas proteínas anormales es una de las metas de las investigaciones en curso.

Como la mayor parte del ATP la generan las mitocondrias a través de la respiración aeróbica, se dice que éstas son las "centrales de energía" de las células. Una célula puede tener desde cientos hasta varios miles de mitocondrias, de acuerdo con su actividad. Las células activas, como las de los músculos, el hígado y los riñones, que utilizan el ATP con gran velocidad, tienen un número elevado de mitocondrias. Las mitocondrias se localizan generalmente en los sitios donde el oxígeno ingresa en la célula o donde se usa el ATP, por ejemplo, entre las proteínas contráctiles de las células musculares.
Una mitocondria está constituido por una membrana mitocondrial externas y una membrana mitocondrial interna, con un pequeño espacio lleno de líquido entre ambas. Las dos membranas son similares en su estructura a la membrana plasmática. La membrana mitocondrial interna contiene una serie de pliegues denominados crestas. La gran cavidad central de una mitocondria, repleta de líquido, es la matriz. Los complejos pliegues de las crestas suministran una superficie extensa para las reacciones químicas que intervienen en la fase aeróbica de respiración celular, reacciones que producen la mayor parte del ATP de la célula. Las enzimas que catalizan estas reacciones están situadas en las crestas y en la matriz mitocondriales.
Como los peroxisomas, las mitocondrias se autorreplican, proceso que tiene lugar durante los intervalos del aumento de la demanda energética por parte de la célula o antes de su división. La síntesis de algunas de las proteínas necesarias para el funcionamiento mitocondrial se produce en los ribosomas de la matriz mitocondrial. Las mitocondrias tienen incluso su propio ADN, en la forma de múltiples copias de moléculas de ADN circular que contienen 37 genes. Estos genes mitocondriales controlan la síntesis de 2 ARN ribosómicos, 22 ARN de transferencia y 14 proteínas que constituyen los componentes mitocondriales.
A pesar de que el núcleo de cada célula somática contiene genes tanto del padre como de la madre, los genes mitocondriales se heredan solo de la madre. La cabeza del espermatozoide (la parte que penetra y fecunda al óvulo) normalmente carece de la mayoría de los orgánulos, como las mitocondrias, los ribosomas, el retículo endoplasmático y el complejo de Golgi, y cualquier mitocondria del espermatozoide que efectivamente pudiese penetrar en el óvulo es destruida rápidamente.
1. Se denomina citoplasma a todo el contenido celular limitado por la membrana plasmática, con excepción del núcleo. Abarca al citosol y los orgánulos.
2. El citosol es la porción líquida del citoplasma y contiene agua, iones, glucosa, aminoácidos, ácidos grasos, proteínas, lípidos, ATP y productos metabólicos. Es el sitio donde se producen muchas reacciones químicas vitales para la célula.
3. Los orgánulos son estructuras especializadas con formas características y que tienen funciones específicas.
4. Los componentes del citoesqueleto, una red de varias clases de filamentos proteicos que se extienden a través del citoplasma, son los microfilamentos, los filamentos intermedios y los microtúbulos. El citoesqueleto suministra un marco estructural a la célula y es responsable de los movimientos celulares.
5. El centrosoma consiste en un par de centríolos y material pericentriolar. El material pericentriolar organiza los microtúbulos en las células que no están en división y el huso mitótico en las las células en división.
6. Los cilios y flagelos, proyecciones móviles de la superficie celular, están formados por los cuerpos basales. Los cilios movilizan los fluidos que se encuentran en la superficie celular; los flagelos mueven la célula entera.
7. Los ribosomas constan de dos subunidades formadas en el núcleo que están compuestas por proteínas y ARN ribosómicos. Son los sitios donde se sintetizan las proteínas.
8. El retículo endoplasmático es una red de membranas que forman sacos aplanados o túbulos; se extiende desde la envoltura nuclear a través del citoplasma.
10. El retículo endoplasmático rugoso está revestido de ribosomas que sintetizan proteínas; éstas ingresan luego en el interior del retículo endoplasmático rugoso para su procesamiento y distribución. El retículo endoplasmático rugoso produce proteínas secretoras, proteínas de membrana y proteínas destinadas a los orgánulos; también forma glucoproteínas, sintetiza fosfolípidos y une las proteínas a los fosfolípidos.
11. El complejo de Golgi está constituido por sacos aplanados llamados cisternas. Las regiones de entrada, medial y de salida del complejo de Golgi contienen diferentes enzimas que le permiten a cada sector modificar, clasificar y agrupar las proteínas para su traslado en vesículas secretoras, en vesículas de membrana o en vesículas de transporte hacia diferentes destinos dentro o fuera de la célula.
12. Los lisosomas son vesículas rodeadas de membrana que contienen enzimas digestivas. Los endosomas, fagosomas y vesículas pinocíticas vierten su contenido en los lisosomas para su degradación posterior. Los lisosomas cumplen funciones en la digestión de los orgánulos viejos (autofagia), en la digestión de las células del huésped (autólisis) y en la digestión extracelular.
13. Los perixosomas contienen oxidasas que oxidan a los aminoácidos, ácidos grasos y sustancias tóxicas; el peróxido de hidrógeno que se produce en estos procesos es inactivado por la enzima catalasa.
14. Las proteasas contenidas en los proteasomas degradan en forma continua las proteínas innecesarias, dañadas o defectuosas mediante su fraccionamiento en péptidos pequeños.
15. La mitocondria consta de una membrana lisa externa, una membrana interna provista de crestas y una cavidad llena de líquido denominada matriz. Estas "centrales de energía" de la célula producen la mayor parte del ATP celular.




El citosol
El citosol (líquido intracelular) es la porción líquida del citoplasma que rodea a los orgánulos y constituye alrededor del 55% del volumen total. A pesar que varía en su composición y consistencia en los distintos sectores de la célula, el citosol está formado en un 70 a 95% por agua, a la que se suman diferentes compuestos que están disueltos o en suspensión.

Entre éstos hay diferentes tipos de iones, glucosa, aminoácidos, ácidos grasos, proteínas, lípidos, ATP y productos de desecho. Ciertas células también presentan diversas moléculas orgánicas que se almacenan en cúmulos. Estos agregados pueden aparecer y desaparecer en diferentes momentos de la vida de una célula. Algunos ejemplos son las "gotas de lípido" que contienen trglicéridos y las partículas de moléculas de glucógeno denominadas gránulos de glucógeno.
El citosol es el lugar donde se producen muchas de las reacciones químicas necesarias para mantener viva a la célula. Por ejemplo, las enzimas del citosol catalizan la glucólisis, una serie de diez reacciones químicas que dan lugar a dos moléculas de ATP a partir de una molécula de glucosa. Otros tipos de reacciones citosólicas aportan los materiales de construcción fundamentales para el mantenimiento y crecimiento de las estructuras celulares.
Orgánulos
Los orgánulos son estructuras especializadas dentro de la célula que tienen unas formas características y que llevan a cabo funciones específicas en el crecimiento, mantenimiento y reproducción celulares. A pesar de la diversidad de reacciones químicas que tienen lugar en una célula en un momento dado, existe muy poca interferencia entre las reacciones ya que están confinadas en diferentes orgánulos.
Cada uno de ellos tiene su propio grupo de enzimas que llevan a cabo reacciones específicas y funcionan como unidades compartimentales para estos procesos bioquímicos determinados. El número y el tipo de orgánulos varían en las diferentes células, de acuerdo con la función que cumplen. A pesar de tener diferentes funciones, los orgánulos cooperan unos con otros para mantener la homeostasis. Si bien el núcleo es un orgánulo grande, será descrito en este artículo debido a su especial importancia en e control del ciclo vital de las células.
El citoesqueleto

El citoesqueleto es una red de filamentos proteicos que se extienden a través del citosol. Tres tipos de filamentos proteicos contribuyen a la estructura del citoesqueleto, así como la de otros orgánulos. En orden creciente de diámetro, estas estructuras son los microfilamentos, los filamentos intermedios y los microtúbulos.
Los microfilamentos, los elementos más delicados del citoesqueleto, están compuestos por la proteína actina y son más abundantes en la periferia de la célula. Tienen dos funciones generales: ayudan a generar el movimiento y proveen soporte mecánico. Con respecto al movimiento, los microfilamentos intervienen en la contracción muscular, la división y la locomoción celulares, como la que se produce en la migración de las células embrionarias durante el desarrollo, la invasión de diferentes tejidos por los glóbulos blancos (leucocitos) para combatir la infección o la migración de células cutáneas en el proceso de curación de las heridas.
Los microfilamentos proporcionan la mayor parte del soporte mecánico que es responsable de la fuerza y la forma básicas de la célula. Estos microfilamentos unen el citoesqueleto a las proteínas integrales de la membrana plasmática, también dan soporte mecánico a las extensiones celulares denominadas microvellosidades, proyecciones en forma de dedo, inmóviles, de la membrana plasmática. Dentro de cada microvellosidad hay un núcleo de microfilamentos paralelos. Puesto que aumentan en gran medida la superficie celular, las microvellosidades son abundantes en las células que están relacionadas con la absorción, como las células epiteliales del intestino delgado.
Como su nombre lo sugiere, los filamentos intermedios son más gruesos que los microfilamentos pero más delgados que los microtúbulos. Varios tipos de proteínas pueden formar parte de los filamentos intermedios, que son excepcionalmente fuertes. Se encuentran en las regiones de la célula sujetas a estrés mecánico, ayudan a estabilizar la ubicación de orgánulos como el núcleo, y contribuyen a que las células se adhieran entre sí.
El componente más grande del citoesqueleto, los microtúbulos, son tubos largos, huecos y no ramificados, compuestos principalmente por la proteína tubulina. El ensamblado de los microtúbulos comienza en un orgánulo llamado centrosoma. Los microtúbulos se desarrollan desde los centrosomas en la periferia de la célula, y contribuyen a dar forma a la célula. También cumplen funciones en el movimiento de ciertos orgánulos como las vesículas secretoras, de los cromosomas durante la división celular y de las proyecciones celulares especializadas, como los cilios y los flagelos.
Centrosoma

El centrosoma, localizado cerca del núcleo, tiene dos componentes: un par de centríolos y material pericentriolar. El centríolo es una estructura cilíndrica compuesta por nueve complejos de tres microtúbulos (tripletes) dispuestos en forma circular. El eje longitudinal de uno de los centríolos está en ángulo recto con el eje longitudinal del otro. Alrededor de los centríolos se encuentra el material centriolar, que contiene cientos de complejos anulares formados por la proteína tubulina. Estos complejos de tubulina son los centros de organización del crecimiento del huso mitótico, que desempeña un papel crítico en la división celular y también en la formación de microtúbulos en las células que no están en división activa. Durante la división celular, los centrosomas se replican de manera que las generaciones sucesivas de células tengan la capacidad de realizar la división celular
Cilios y flagelos

Los microtúbulos con los componentes predominantes de los cilios y flagelos, protecciones móviles de la superficie celular. Los cilios son apéndices numerosos, cortos, similares a pelos, que se extienden desde la superficie de la célula. Cada cilio contiene un núcleo de 20 microtúbulos rodeado por la membrana plasmática. Los microtúbulos están dispuestos de tal manera que un par central es rodeado por nueve complejos de dos microtúbulos fusionados (dobletes). Cada cilio permanece unido a un cuerpo basal que se halla por debajo de la superficie de la membrana plasmática. Un cuerpo basal es similar en su estructura a un centríolo y cumple funciones en el ensamblado de los cilios y flagelos.
Los cilios realizan un movimiento similar al de un remo; es relativamente rígido durante el movimiento de fuerza (el remo introduciéndose en el agua), pero es más flexible durante el movimiento de recuperación (el remo moviéndose por arriba del agua y preparándose para una nueva remada). Los movimientos coordinados de muchos cilios en la superficie de una célula dan lugar a un movimiento sostenido del líquido a lo largo de la superficie celular.

Muchas células del tracto respiratorio, por ejemplo, tiene cientos de cilios que ayudan a barrer fuera de los pulmones las partículas extrañas atrapadas en el moco. En la fibrosis quística, la densidad anormal de las secreciones mucosas interfiere con la acción de los cilios y, por ende, con las funciones normales del tracto respiratorio. El movimiento de los cilios también es paralizado por la nicotina del humo del cigarrillo. Por esta razón, los fumadores tosen con frecuencia para eliminar las partículas extrañas de sus vías respiratorias. Las células que revisten las trompas uterinas (de Falopio) también tienen cilios que desplazan al óvulo hacia el útero, y las mujeres que fuman tienen un riesgo mayor de embarazo ectópico (fuera del útero).
Los flagelos son similares en su estructura a los cilios, pero suelen ser mucho más largos. Los flagelos generalmente mueven una célula entera. In flagelo genera movimiento hacia adelante a lo largo de su eje por el movimiento rápido que sigue un patrón ondulante. El único ejemplo de flagelos en el cuerpo humano e la cola de los espermatozoides, que propulsa a estas células hacia su encuentro con el óvulo.
Ribosomas

Los ribosomas son los sitios de síntesis proteica. El nombre de estos pequeños orgánulos refleja su alto contenido de un tipo especial de ácido ribonucleico, el ARN ribosómico (ARNr), aunque también puede incluir a más de 50 proteínas. Estructuralmente, un ribosoma está constituido por subunidades, una de las cuales tiene la mitad del tamaño de la otra. Las subunidades mayor y menor se forman por separado en el nucléolo, un cuerpo esférico que está dentro del núcleo. Una vez sintetizadas, las subunidades mayor y menor abandonan el núcleo y se unen en el citoplasma.
Algunos ribosomas están adheridos a la superficie externa de la membrana nuclear y a una membrana extensamente plegada denominada retículo endoplasmático. Estos ribosomas sintetizan las proteínas que tienen como destino orgánulos específicos, su inserción en la membrana plasmática o su salida de la célula. Otros ribosomas son "libres" o no adheridos a ninguna otra estructura citoplasmática. Los ribosomas libres sintetizan proteínas que serán utilizadas en el citosol. Los ribosomas también están localizados dentro de las mitocondrias, donde sintetizan proteínas mitocondriales.
Retículo endoplasmático

El retículo endoplsamático es una red de membranas en forma de sacos aplanados o túbulos. El retículo endoplasmático se extiende desde la membrana o envoltura nuclear (membrana que rodea al núcleo), con la cual se conecta hacia todo el citoplasma. El retículo endoplasmático es tan amplio que constituye casi la mitad de las superficies membranosas dentro del citoplasma de la mayoría de las células.
Las células contienen dos tipos distintos de retículo endoplasmático, que difieren tanto en su estructura como en su función. El retículo endoplasmático rugoso es una estructura que continúa con la membrana nuclear y generalmente se presenta plegado a una serie de sacos aplanados. La superficie externa del retículo endoplasmático rugoso está revestida de ribosomas, los sitios de síntesis proteica. Las proteínas sintetizadas por los ribosomas penetran en los espacios que hay en el interior del retículo endoplasmático rugoso para su procesamiento y distribución. En algunos casos, ciertas enzimas unen proteínas con hidratos de carbono para formar glucoproteínas. En otros casos, las enzimas unen proteínas con fosfolípidos, también sintetizados en el retículo endoplasmático rugoso. Estás moléculas pueden incorporarse a las membranas de los orgánulos, insertarse en la membrana plasmática o secretarse por exocitosis. De tal modo que el retículo endoplasmático rugoso produce proteínas secretoras, proteínas de membrana y muchas proteínas de los orgánulos.
El retículo endoplasmático liso se extiende desde el retículo endoplasmático rugoso para formar una red de túbulos membranosos. A diferencia del R.E. rugoso, el retículo endoplasmático liso carece de ribosomas en la superficie externa de sus membranas. Sin embargo, contiene enzimas especiales que pueden hacerlo funcionalmente más diverso que el retículo endoplasmático rugoso. La ausencia de ribosomas impide la síntesis de proteínas, pero sí puede sintetizar ácidos grasos y esteroides, como los estrógenos y la testosterona. En las células hepáticas, las enzimas del retículo endoplasmático liso facilitan la liberación de la glucosa hacia el torrente sanguíneo y contribuyen a inactivar o detoxificar los fármacos liposolubles o las sustancias potencialmente nocivas, como el alcohol, los pesticidas y los carcinógenos (agentes causantes de cáncer). En las células del hígado, del riñón y del intestino, una enzima del retículo endoplasmático liso separa el grupo fosfato de la glucosa-6-fosfato, lo cual permite que se libere la glucosa y pueda ingresar en la sangre. En las células musculares, los iones calcio que estimulan la contracción son liberados desde el retículo sarcoplasmático, una forma del retículo endoplasmático liso.
Retículo endoplasmático liso y la tolerancia a los fármacos
Una de las funciones del retículo endoplasmático liso es detoxificar ciertos fármacos. Quienes consumen de forma repetido algunos fármacos, como el sedante fenobarbital, desarrollan cambios en el retículo endoplasmático listo de las células hepáticas. La administración prolongada de fenobarbital trae como resultado la tolerancia al fármaco; la misma dosis deja de producir el mismo grado de sedación. Con la exposición reiterada al fármaco, la cantidad del retículo endoplasmático liso y sus enzimas aumentan para proteger a la célula de sus efectos tóxicos. Como la cantidad de retículo endoplasmático liso aumenta, son necesarias dosis cada vez más altas del fármaco para alcanzar el grado original del efecto.
Complejo de Golgi

La mayor parte de las proteínas sintetizadas en los ribosomas adheridos al retículo endoplasmático rugoso son por último transportadas a otras regiones de la célula. El primer paso en la vía de transporte es un orgánulo denominado complejo de Golgi. Consta de 2 a 20 cisternas, pequeños sacos aplanados membranosos, de bordes salientes, que parecen una pila de pitas (pan árabe). Las cisternas suelen ser curvadas y le dan forma al complejo de Golgi un aspecto de copa. Casi todas las células tienen varios complejos de Golgi, y éstos son más numerosos en las células que secretan proteínas, una clave para entender el papel de este orgánulo en la célula.
Las cisternas en los extremos opuestos de un complejo de Golgi difieren entre sí en su forma, su tamaño y su actividad enzimática. La entrada convexa o cara cis, mira hacia el retículo endoplasmático rugoso. La salida cóncava o cara trans, mira hacia la membrana plasmática. Los sacos que se hallan entre las caras de entrada y de salida se denominan cisternas mediales. Las vesículas de transporte provenientes del retículo endoplasmático se fusionan para formar la cara de entrada. Se piensa que las cisternas mediales y luego en cisternas de salida.
Las diferentes enzimas presentes en las regiones de entrada, medial y de salida del complejo de Golgi permiten que cada una de estas áreas pueda modificar, ordenar y envolver a las proteínas para su transporte a diferentes destinos. La cara de entrada recibe y modifica las proteínas producidas por el retículo endoplasmático rugoso. Las cisternas mediales agregan hidratos de carbono a las proteínas para formar glucoproteínas y lípidos para formar lipoproteínas. La cara de salida modifica aún más las moléculas y luego las selecciona y reúne para el transporte hacia su destino final.
Las proteínas que llegan al complejo de Golgi, lo atraviesan y salen de éste mediante la maduración de las cisternas y los intercambios que ocurren por las vesículas de transferencia.

1. Las proteínas sintetizadas por los ribosomas del retículo endoplasmático rugoso son rodeadas por una porción de la membrana del retículo endoplasmático, que posteriormente brota de la superficie de la membrana para formar las vesículas de transporte.
2. Las vesículas de transporte se dirigen hacia la cara de entrada del complejo de Golgi.
3. La fusión de varias vesículas de transporte crea una cara de entrada en el complejo de Golgi y libera proteínas dentro de su luz (espacio).
4. Las proteínas se mueven desde la cara de entrada hacia una o más cisternas mediales. Las enzimas de estas cisternas modifican las proteínas para formar glucoproteínas, glucolípidos, y lipoproteínas. Las vesículas de transferencia que surgen de los bordes de las cisternas llevan enzimas específicas de regreso a la cara de entrada y transportan algunas proteínas parcialmente modificadas hacia la cara de salida.
5. Los productos de las cisternas mediales se mueven hacia la luz de la cara de salida.
6. Dentro de la cara de salida, los productos sufren más modificaciones, se clasifican y rodean de membrana y son empaquetados.
7. Algunas de las proteínas procesadas abandonan la cara de saluda y quedan almacenadas en vesículas secretoras. Estas vesículas llevan las proteínas hacia la membrana plasmática, donde se liberan por exocitosis hacia el líquido extracelular. Ciertas células pancreáticas, por ejemplo, liberan la hormona insulina de esta forma.
8. Otras proteínas procesadas abandonan la cara de salida en vesículas de membrana que entregan su contenido a la membrana plasmática para su incorporación dentro de la propia membrana. De esta manera, el complejo de Golgi agrega unos nuevos segmentos a la membrana plasmática a medida que los preexistentes se pierden y modifica el número y la distribución de las moléculas de la membrana.
9. Finalmente, algunas proteínas procesadas abandonan la cara de salida de las vesículas de transporte que las llevan hacia otro destino en la célula. Por ejemplo, las vesículas de transporte llevan las enzimas digestivas hacia los lisosomas.
Lisosomas

Los lisosomas son vesículas rodeadas de membrana que se forman en el complejo de Golgi. En su interior hay más de 60 tipos de poderosas enzimas digestivas e hidrolíticas que pueden digerir una gran variedad de moléculas una vez que los lisosomas se fusionaron con las vesículas formadas durante la endocitosis. Como las enzimas lisosómicas funcionan mejor en un pH ácido, la membrana lisosómica contiene bombas de transporte activas que importan iones hidrógeno (H+). De esta manera, el interior de los lisosomas tiene un pH de 5, que es 100 veces más ácido que el pH del citosol (pH 7). La membrana lisosómica también presenta transportadores que acarrean los productos finales de la digestión, como la glucosa, los ácidos grasos y los aminoácidos, hacia el citosol.
Las enzimas lisosómicas intervienen, así mismo, en el reciclado de las estructuras celulares deterioradas. Un lisosoma puede ingerir otro orgánulo, digerirlo y luego devolver los componentes digeridos al citosol para su nuevo uso, proceso que se denomina autofagia. Durante la autofagia, el orgánulo que ha de ser digerido es rodeado por una membrana que deriva del retículo endoplasmático para crear la vesícula denominada autofagosoma, que luego se fusiona con un lisosoma. De tal modo, una célula hepática humana recicla alrededor de la mitad de su contenido citoplasmático cada semana. Las enzimas lisosómicas también pueden destruir la célula entera mediante el proceso de autólisis. La autólisis tiene lugar en algunas situaciones patológicas y es también responsable del deterioro de los tejidos después de la muerte.
La mayor parte de las enzimas lisosómicas actúan dentro de la célula. Sin embargo, algunas participan en la digestión extracelular. Un ejemplo se da durante la fecundación. La cabeza del espermatozoide libera enzimas lisosómicas que lo ayudan a introducirse en el óvulo, por la disolución de su cubierta protectora en el proceso llamado reacción acrosómica.
Peroxisomas
Otro grupo de orgánulos similares en su estructura a los lisosomas, pero más pequeños, son los peroxisomas. Contienen varias oxidasas, enzimas que pueden oxidar (eliminar átomos de hidrógeno) de diversas sustancias orgánicas. Por ejemplo, los aminoácidos y los ácidos grasos pueden ser oxidados en los peroxisomas como parte del metabolismo normal. Además, las enzimas de los peroxisomas oxidan sustancias tóxicas, como el alcohol. Por ello son muy abundantes en el hígado, donde tiene lugar la detoxificación del alcohol y otras sustancias nocivas. Un producto intermedio de las reacciones de oxidación es el peróxido de hidrógeno (H2O2), un compuesto potencialmente tóxico. Sin embargo, los peroxisomas también contienen la enzima catalasa, que descompone al H2O2. Como la producción y la degradación del H2O2 tiene lugar dentro de la misma organela, los peroxisomas protegen a otras partes de la célula de los efectos tóxicos del H2O2. Los peroxisomas nuevos se forman a partir de los ya existentes.
Proteasomas
Como ya mencionó, los lisosomas degradan las proteínas que reciben por medio de las vesículas. Las proteínas citosólicas también requieren su eliminación en algún momento del ciclo vital de una célula. La destrucción permanente de las proteínas no necesarias, dañadas o defectuosas es una función que está a cargo de pequeñas estructuras con forma de tonel denominadas proteasomas. Las proteínas que forman parte de las vías metabólicas, por ejemplo, son degradas después de haber cumplido su función. Esta destrucción proteica participa en la retroalimentación negativa ya que establece un alto en la vía una vez que se obtuvo la respuesta adecuada. Una célula típica del organismo contiene muchos cientos de proteasomas, tanto en el citosol como en el núcleo. Los proteasomas reciben su nombre debido a su alto contenido de proteasas, enzimas que pueden cortar a las proteínas en péptidos pequeños. Una vez que las enzimas de un proteasoma hidrolizaron a una proteína en segmentos más pequeños, otras enzimas pueden luego descomponer a esos péptidos en aminoácidos, los cuales se reciclan para formar nuevas proteínas.
Algunas enfermedades son el resultado de la incapacidad de los proteasomas de degradar proteínas anormales. Por ejemplo, se acumulan grupos de proteínas con defectos de plegamiento en las células cerebrales en los pacientes con enfermedad de Parkinson y enfermedad de Alzheimer. El descubrimiento de por qué los proteasomas no pueden degradar a estas proteínas anormales es una de las metas de las investigaciones en curso.
Mitocondrias

Como la mayor parte del ATP la generan las mitocondrias a través de la respiración aeróbica, se dice que éstas son las "centrales de energía" de las células. Una célula puede tener desde cientos hasta varios miles de mitocondrias, de acuerdo con su actividad. Las células activas, como las de los músculos, el hígado y los riñones, que utilizan el ATP con gran velocidad, tienen un número elevado de mitocondrias. Las mitocondrias se localizan generalmente en los sitios donde el oxígeno ingresa en la célula o donde se usa el ATP, por ejemplo, entre las proteínas contráctiles de las células musculares.
Una mitocondria está constituido por una membrana mitocondrial externas y una membrana mitocondrial interna, con un pequeño espacio lleno de líquido entre ambas. Las dos membranas son similares en su estructura a la membrana plasmática. La membrana mitocondrial interna contiene una serie de pliegues denominados crestas. La gran cavidad central de una mitocondria, repleta de líquido, es la matriz. Los complejos pliegues de las crestas suministran una superficie extensa para las reacciones químicas que intervienen en la fase aeróbica de respiración celular, reacciones que producen la mayor parte del ATP de la célula. Las enzimas que catalizan estas reacciones están situadas en las crestas y en la matriz mitocondriales.
Como los peroxisomas, las mitocondrias se autorreplican, proceso que tiene lugar durante los intervalos del aumento de la demanda energética por parte de la célula o antes de su división. La síntesis de algunas de las proteínas necesarias para el funcionamiento mitocondrial se produce en los ribosomas de la matriz mitocondrial. Las mitocondrias tienen incluso su propio ADN, en la forma de múltiples copias de moléculas de ADN circular que contienen 37 genes. Estos genes mitocondriales controlan la síntesis de 2 ARN ribosómicos, 22 ARN de transferencia y 14 proteínas que constituyen los componentes mitocondriales.
A pesar de que el núcleo de cada célula somática contiene genes tanto del padre como de la madre, los genes mitocondriales se heredan solo de la madre. La cabeza del espermatozoide (la parte que penetra y fecunda al óvulo) normalmente carece de la mayoría de los orgánulos, como las mitocondrias, los ribosomas, el retículo endoplasmático y el complejo de Golgi, y cualquier mitocondria del espermatozoide que efectivamente pudiese penetrar en el óvulo es destruida rápidamente.
Características y datos específicos del citoplasma y los orgánulos
1. Se denomina citoplasma a todo el contenido celular limitado por la membrana plasmática, con excepción del núcleo. Abarca al citosol y los orgánulos.
2. El citosol es la porción líquida del citoplasma y contiene agua, iones, glucosa, aminoácidos, ácidos grasos, proteínas, lípidos, ATP y productos metabólicos. Es el sitio donde se producen muchas reacciones químicas vitales para la célula.
3. Los orgánulos son estructuras especializadas con formas características y que tienen funciones específicas.
4. Los componentes del citoesqueleto, una red de varias clases de filamentos proteicos que se extienden a través del citoplasma, son los microfilamentos, los filamentos intermedios y los microtúbulos. El citoesqueleto suministra un marco estructural a la célula y es responsable de los movimientos celulares.
5. El centrosoma consiste en un par de centríolos y material pericentriolar. El material pericentriolar organiza los microtúbulos en las células que no están en división y el huso mitótico en las las células en división.
6. Los cilios y flagelos, proyecciones móviles de la superficie celular, están formados por los cuerpos basales. Los cilios movilizan los fluidos que se encuentran en la superficie celular; los flagelos mueven la célula entera.
7. Los ribosomas constan de dos subunidades formadas en el núcleo que están compuestas por proteínas y ARN ribosómicos. Son los sitios donde se sintetizan las proteínas.
8. El retículo endoplasmático es una red de membranas que forman sacos aplanados o túbulos; se extiende desde la envoltura nuclear a través del citoplasma.
10. El retículo endoplasmático rugoso está revestido de ribosomas que sintetizan proteínas; éstas ingresan luego en el interior del retículo endoplasmático rugoso para su procesamiento y distribución. El retículo endoplasmático rugoso produce proteínas secretoras, proteínas de membrana y proteínas destinadas a los orgánulos; también forma glucoproteínas, sintetiza fosfolípidos y une las proteínas a los fosfolípidos.
11. El complejo de Golgi está constituido por sacos aplanados llamados cisternas. Las regiones de entrada, medial y de salida del complejo de Golgi contienen diferentes enzimas que le permiten a cada sector modificar, clasificar y agrupar las proteínas para su traslado en vesículas secretoras, en vesículas de membrana o en vesículas de transporte hacia diferentes destinos dentro o fuera de la célula.
12. Los lisosomas son vesículas rodeadas de membrana que contienen enzimas digestivas. Los endosomas, fagosomas y vesículas pinocíticas vierten su contenido en los lisosomas para su degradación posterior. Los lisosomas cumplen funciones en la digestión de los orgánulos viejos (autofagia), en la digestión de las células del huésped (autólisis) y en la digestión extracelular.
13. Los perixosomas contienen oxidasas que oxidan a los aminoácidos, ácidos grasos y sustancias tóxicas; el peróxido de hidrógeno que se produce en estos procesos es inactivado por la enzima catalasa.
14. Las proteasas contenidas en los proteasomas degradan en forma continua las proteínas innecesarias, dañadas o defectuosas mediante su fraccionamiento en péptidos pequeños.
15. La mitocondria consta de una membrana lisa externa, una membrana interna provista de crestas y una cavidad llena de líquido denominada matriz. Estas "centrales de energía" de la célula producen la mayor parte del ATP celular.
¿Tienes algo que decir sobre este artículo? Compártelo con nosotros en:



Comentarios
Publicar un comentario